|
|
01/05/1997 La
Recherche
BERNARD
DUTRILLAUX
Institut Curie, UMR 147 du CNRS et
Département de radiobiologie et radiopathologie DSV/CEA.
FLORENCE RICHARD
Institut Curie, UMR 147 du
CNRS et université Versailles/ St-Quentin/Yvelines.
N.B.
Cet article complète un précédent article de Bernard
Dutrillaux, paru dans notre numéro spécial sur l'évolution
(mars 1997). Lequel article peut également être lu sur notre
site Web.
 L'évolution des techniques
L'évolution des techniques
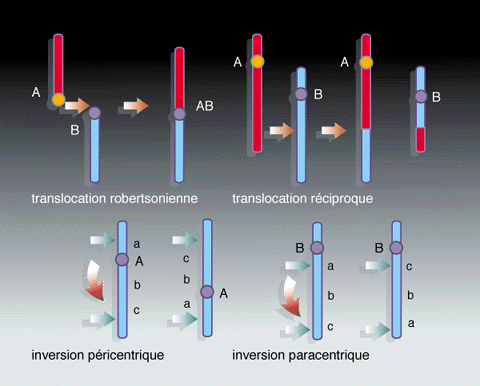
 Voici quatre types
de remaniements chromosomiques Voici quatre types
de remaniements chromosomiques
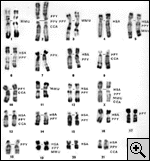 Voici le caryotype
présumé de l'ancêtre commun à l'homme, Voici le caryotype
présumé de l'ancêtre commun à l'homme,
*LE CARYOTYPE
est l'ensemble des chromosomes d'une cellule normale de
l'individu. Chez l'homme, il comprend 46 chromosomes, dont
deux sexuels.
*LE POLYMORPHISME
est la qualité d'une espèce ou d'une population qui se
présente sous plusieurs formes. D'un individu à l'autre, on
observe notamment des variations dans les gènes ou parfois les
chromosomes, qui retentissent alors sur le phénotype*.
*HÉTÉROZYGOTE
De hetero
(différent) et zygote (oeuf fécondé). Employé ici au
sens d'un individu dont les deux allèles (copies provenant de
l'un et l'autre parent) d'un même gène sont différents.
*LE PHÉNOTYPE
est l'ensemble des
caractères morphologiques et biologiques d'un individu,
déterminés par ses gènes (génotype).
*LE
CENTROMÈRE
est la région du chromosome qui relie les
deux chromatides, leur donnant leur forme en X ou en V. Il se
situe dans une région riche en ADN répété. La fonction
principale du centromère est l'attachement du chromosome au
fuseau mitotique qui se forme lors de la division cellulaire,
permettant la migration des chromosomes vers chaque pôle de la
cellule.
*Un chromosome est ACROCENTRIQUE quand son
centromère est situé à une extrémité (forme en V). Un
chromosome en forme de X est appelé MÉTACENTRIQUE.
*CHIMPANZÉS BLANCS ET NOIRS
Tous les
chimpanzés ont le pelage foncé. Toutefois, la pigmentation
cutanée varie, ce qui permet de distinguer des chimpanzés à
face blanche et d'autres à face noire.
| |
|
|
|
> ARTICLES
>
CYTOGÉNÉTIQUE
L'analyse des chromosomes permet de
réécrire l'histoire des primates
Notre nouvel arbre de famille
La
comparaison systématique des chromosomes de quelque cent vingt
espèces de primates permet de reconstituer de manière précise
l'arbre généalogique de notre grande famille. Jamais sans
doute la place de l'espèce humaine dans le monde vivant n'aura
été identifiée comme aussi banale. L'analyse réserve aussi
quelques surprises.
L'analyse des chromosomes
d'individus appartenant à une même espèce montre généralement
qu'ils possèdent des caryotypes*
semblables. Toutefois , un polymorphisme*
chromosomique plus ou moins grand peut exister, ce qui traduit
une dynamique montrant que la notion d'espèce stable, telle
qu'elle est souvent comprise, est un peu simpliste. Une espèce
est un ensemble d'individus qui partagent plus ou moins
totalement un ensemble de caractères héréditaires. Cet
ensemble de caractères n'est pas exactement celui d'hier et
sera encore modifié demain. L'ensemble de ces modifications
n'est pas partagé par toute la population, de sorte que
s'établit un polymorphisme, qui pourra être plus ou moins
marqué selon les périodes et les régions de distribution de
l'espèce. Il n'en reste pas moins que, dans la grande majorité
des cas, il est possible de définir le caryotype d'une espèce,
car ses variations sont le plus souvent modérées. Ainsi, chez
l'homme, qui est le mammifère le plus étudié, 995 individus
sur mille possèdent le même caryotype à 46 chromosomes.
Environ un caryotype sur mille possède 45 chromosomes, à la
suite d'une translocation robertsonienne (fig. 1), et un
caryotype sur mille un autre remaniement chromosomique
équilibré (sans effet pathologique décelable).
Ces particularités sont presque toujours observées à
l'état hétérozygote*, c'est-à-dire qu'elles ne concernent
qu'un seul chromosome d'une paire. Les autres formules
aberrantes sont déséquilibrées, donc associées à une
pathologie, et, n'étant pas transmises à la descendance,
n'interviennent pas dans le polymorphisme à proprement parler.
Lorsque les caryotypes représentatifs de deux espèces
sont comparés, des différences sont généralement constatées.
Elles portent sur le nombre ou la morphologie des chromosomes,
ou les deux en même temps. Ainsi, chez les primates, les
formules chromosomiques les plus basses sont observées chez
les lépilemurs de Madagascar (2N = 20), les plus élevées chez
les tarsiers d'Asie (2N = 84).
Les primates comprennent actuellement environ deux
cents espèces. Dans nos travaux, nous avons analysé le
caryotype de près de cent vingt d'entre elles. Commençons par
un bref rappel de la systématique. L'ordre des primates
comprend deux sous-ordres, celui des prosimiens et celui des
simiens. Le sous-ordre des simiens comprend deux infra-ordres.
Celui des catarhiniens - où l'homme se situe - est ainsi nommé
parce qu'ils ont des narines rapprochées et celui des
platyrhiniens, dont les narines sont écartées et dont les
espèces sont toutes sur le continent américain. Le sous-ordre
des prosimiens contient quatre infra-ordres, les tupaiiformes,
les lémuriens, les lorisiformes et les tarsiers. Les deux
cents espèces de primates se distribuent à peu près à parts
égales entre catarhiniens, platyrhiniens et prosimiens. Ces
derniers se localisent pour une grande part à Madagascar
(lémurs) et au sud des continents africain et asiatique.
La comparaison des caryotypes permet quelques
conclusions simples.
D'abord, beaucoup d'espèces considérées comme proches
par les systématiciens, c'est-à-dire appartenant au même
genre, voire à la même famille, possèdent des caryotypes
proches, dont beaucoup de chromosomes sont identiques.
On peut en déduire que deux paramètres relativement
indépendants ont évolué parallèlement : d'une part les
modifications chromosomiques, et d'autre part les mutations
des gènes responsables des traits morphologiques sur lesquels
la systématique classique se fonde. Mais il y a des
exceptions. Ainsi, des espèces aux phénotypes* nettement
différents, comme les macaques, babouins, géladas, drills et
mandrills, peuvent posséder des caryotypes identiques ou très
semblables. A l'inverse, des espèces proches, comme les divers
cercopithèques arboricoles, paraissent morphologiquement
proches, mais ont des caryotypes assez différents. En dépit de
ces exceptions, plus les espèces sont éloignées, plus il est
difficile de comparer leurs chromosomes directement.
Il nous a donc fallu procéder par une approche
progressive de comparaison d'espèces proches, afin d'en
déduire les éléments communs, qui représentent en général les
caractères ancestraux. Par exemple, le fait que le chimpanzé,
le gorille et l'orang-outan possèdent un chromosome identique
au chromosome 6 humain indique que leur ancêtre commun
possédait déjà ce chromosome. En suivant ce principe, dit de
parcimonie car il adopte l'hypothèse la plus simple (la plus
économe), il est possible de reconstituer avec une bonne
précision le caryotype ancestral d'un groupe d'espèces
proches, comme celui de l'homme ( Hominidae ) et des
grands singes ( Pongidae ). On peut en faire autant
pour les Cercopithecidae , beaucoup plus nombreux en
espèces.
Il devient alors possible de comparer les deux
caryotypes ainsi reconstitués, ce qui s'avère très riche en
informations. Ainsi, lorsque les deux reconstitutions,
indépendantes, ont amené à désigner le même chromosome comme
ancestral, il devient certain qu'il l'était effectivement. De
plus, le choix d'un chromosome ancestral n'étant pas toujours
univoque dans un groupe donné, lorsque l'une des solutions
possibles se retrouve dans deux groupes indépendants, cette
solution devient très probable.
Nous avons donc progressivement reconstitué le
caryotype ancestral des catarhiniens, puis celui des
platyrhiniens et des prosimiens, et il a été très satisfaisant
de constater qu'ils partageaient de nombreux chromosomes
identiques ou très semblables. Cette constatation avait valeur
de confirmation, puisqu'il est hautement improbable de
reconstituer par hasard de mêmes chromosomes indépendamment.
La comparaison de ces trois caryotypes ancestraux permet alors
de franchir une nouvelle étape, qui est celle de la
reconstitution du caryotype ancestral de l'ensemble des
primates.
Pour valider ce caryotype, il était nécessaire de le
comparer à celui d'autres animaux. C'est ainsi que nous avons
effectué le même travail de reconstitution pour les
carnivores. A nouveau, leur caryotype ancestral s'est avéré
très semblable à celui reconstitué pour les primates, ce qui
nous a permis d'approcher la reconstitution du caryotype
ancestral des mammifères placentaires.
Parmi les nombreux caryotypes d'espèces rattachées à
d'autres ordres, nous avons alors recherché si certains
chromosomes ancestraux s'y retrouvaient. Cela fut le cas,
montrant au passage que certaines espèces avaient conservé de
nombreux chromosomes ancestraux, alors que d'autres n'en
possédaient plus.
Ainsi, chez les rongeurs, la famille des écureuils (
Sciuridae ) a conservé des chromosomes peu modifiés, donc
faciles à comparer à ceux des primates. A l'inverse, la
famille des Muridae , à laquelle se rattachent les rats
et les souris, a des chromosomes très différenciés. Ainsi, en
se basant sur cette différenciation, la souris possède l'un
des caryotypes les plus « évolués », beaucoup plus évolué, par
rapport au caryotype de l'ancêtre, que celui de l'homme, par
exemple. Ce n'est pas anecdotique, car la souris est le
mammifère le plus étudié avec l'homme. Son génome est donc
bien connu, et les extrapolations à l'homme sont d'un grand
intérêt.
Malheureusement, le hasard a fait que l'animal le plus
usité pour l'expérimentation soit aussi l'un de ceux dont le
génome est le plus éloigné de celui de notre propre espèce...
Le surcoût d'un tel choix serait considérable, s'il n'était
compensé par d'autres avantages, mais cela complique
sérieusement la tâche des biologistes.
D'autres animaux ont conservé des caryotypes primitifs.
C'est le cas du lièvre et du lapin dans l'ordre des
lagomorphes. C'est aussi le cas du tatou, dans l'ordre des
édentés. Finalement, toutes ces intercomparaisons nous ont
permis de reconstruire un caryotype ancestral commun, avec une
précision probablement assez bonne. On peut en effet estimer
que 80 % du caryotype du dernier ancêtre commun des mammifères
placentaires, qui a vécu il y a une centaine de millions
d'années, est connu, avec une précision qui va très au-delà
des notions que nous avons sur sa morphologie.
A titre d'exemple nous présentons le caryotype présumé
du dernier ancêtre commun aux Hominidae , Pongidae
et Cercopithecidae (fig. 2). Seule ambiguïté
persistante, le chromosome 3 de cet ancêtre était soit comme
chez l'orang-outan actuel, soit comme chez le macaque.
L'animal, dont le caryotype est représenté, a vécu il y a 30 à
50 millions d'années.
Certains principes élémentaires de la reconstitution
phylogénétique ayant été évoqués dans La Recherche de
mars 1997, il n'est pas utile de les reprendre en détail ici.
Rappelons toutefois que, du fait que les chromosomes possèdent
des structures en bandes, il est possible de reconstituer
précisément les réarrangements qu'ils subissent. Il est de
même possible, lorsqu'un chromosome subit plusieurs
modifications, d'identifier d'éventuelles étapes
intermédiaires, et donc d'ordonner une suite d'événements. La
reconstitution des étapes ancestrales, telle que nous venons
de l'évoquer plus haut, permet ensuite de préciser l'origine,
et donc de fixer ce qui est ancestral et ce qui en est dérivé.
Ceci est un énorme avantage par rapport à beaucoup d'autres
caractères, morphologiques en particulier, dont il est souvent
difficile de déterminer la nature ancestrale ou dérivée.
La reconstitution de la phylogénie chromosomique a
bénéficié de deux autres avantages, directement liés à la
mécanique évolutive. D'abord, le taux de réarrangements
séparant le caryotype d'espèces proches dépasse rarement la
dizaine, ce qui est très informatif et rend l'interprétation
possible (un grand nombre de réarrangements rendrait
l'information trop complexe).
Ensuite, la plupart des réarrangements sont soit
intrachromosomiques (inversions et fissions), soit, quand ils
sont interchromosomiques, relativement simples (translocations
robertsoniennes). Une accumulation d'autres remaniements
rendrait le décryptage trop complexe. C'est ce qui est arrivé,
par exemple chez les muridés, dont nous avons déjà parlé, et,
plus proches de nous, chez les gibbons. Il nous a été
impossible, pour cette raison, de situer précisément les
gibbons parmi les autres primates. Ces reconstitutions
chromosomiques nous ont donc permis de proposer l'arbre «
généalogique » représenté page suivante.
Cet arbre indique pour commencer que l'évolution des
primates a été très vite divergente. A vrai dire, il n'est
même pas possible de proposer l'existence d'un tronc commun,
puisqu'on assiste d'emblée à une bifurcation entre simiens et
prosimiens ancestraux. Cette absence de remaniement
chromosomique commun n'exclut pas, toutefois, l'existence d'un
tronc commun où seules des mutations géniques seraient
survenues.
Les prosimiens ont aussi une divergence très précoce,
entre espèces malgaches et afro-asiatiques. Le groupe malgache
(lémuriformes) comprend plusieurs familles, parmi lesquelles
les aye-aye ( Daubentoniidae , DMA) se sont séparés
tôt. Les autres familles, Cheirogaleidae ,
Lemuridae , Indriidae et Lepilemuridae se
sont ensuite toutes isolées à partir d'une même population
ancestrale (LEM).
Le groupe afro-asiatique (lorisiformes, LOR) comprend
deux familles, les Lorisidae (à droite) et les
Galagidae . Celles-ci partagent un long tronc commun,
et se trouvent ensuite bien séparées. Nous n'avons pu obtenir
d'informations permettant de situer les tarsiers. Ils ont
vraisemblablement subi une évolution chromosomique avec de
nombreuses fissions et translocations, rendant leur caryotype
très difficile à comparer à celui des autres primates. Quant
aux tupaiiformes, leur caryotype est relativement simple. Il
n'a pas été pris en compte car l'appartenance des tupaiiformes
à l'ordre des primates est incertaine.
On remarquera que la très grande majorité des
modifications chromosomiques survenues chez les prosimiens
sont des translocations robertsoniennes (indiquées par des
carrés clairs).
A l'origine de la plupart des branches menant aux
espèces, des cercles ont été indiqués dans lesquels les
remaniements sont partagés diversement. Ceci traduit la
survenue d'une évolution en réseau, dont nous avons détaillé
le principe dans le numéro de mars de La Recherche . La
seule évolution franchement dichotomique (par bipartition) est
observée dans le groupe des Lepilemuridae , dont le
nombre chromosomique a été très réduit par la survenue de
nombreuses fusions.
Les simiens partagent eux aussi un tronc très court,
avant de se séparer en platyrhiniens et catarhiniens.
Chez les platyrhiniens, les deux grandes familles se
trouvent également bien séparées. Les Callitrichidae ,
qui regroupent les tamarins et les ouistitis, partagent un
tronc commun avant de se séparer par une évolution en réseau.
L'évolution des Cebidae , qui regroupent les autres,
semble plus complexe, et notre reconstitution est encore
inachevée. C'est probablement dans cette famille que les
révisions systématiques les plus grandes doivent être
apportées. Les positions respectives des singes capucins (
Cebus ), des singes écureuils ( Saimiri ) et des
singes araignées ( Ateles ), entre autres, longtemps
imprécises, se trouvent maintenant indiquées par notre schéma.
L'ensemble des catarhiniens partage un long tronc
commun, avant la bifurcation séparant la nombreuse famille des
Cercopithecidae . Un long tronc commun est à nouveau
partagé par les trois sous-familles, Colobinae ,
Papioninae et Cercopithecinae, issues d'une même
population commune. L'évolution des Papioninae (macaca,
papio, cercocebus, mandrillus) se caractérise par la stabilité
du caryotype, resté très proche de celui de l'ancêtre commun
des Cercopithecidae . Ce groupe offre un démenti aux
hypothèses qui supposaient que la spéciation passe
nécessairement par des modifications chromosomiques.
L'évolution des Cercopithecinae est plus remarquable.
Plus de cinquante réarrangements séparent les caryotypes d'un
nombre d'espèces comparable à celui des Papioninae .
Etant issues d'une même population ancestrale, ces deux
sous-familles ont donc accumulé les modifications
chromosomiques à des vitesses très différentes. La survenue de
ces réarrangements ne peut donc constituer une horloge
biologique. Autre particularité, l'évolution des
Cercopithecinae a séparé deux populations ayant ensuite
évolué en réseaux. De l'une de ces populations sont issues les
espèces terrestres, de l'autre la plupart des espèces
arboricoles. On remarquera, en outre, l'accumulation de
fissions, indiquées par des flèches, qui contribuent chacune à
augmenter le nombre de chromosomes. C'est ainsi que certaines
espèces, comme les mones, possèdent plus de 70 chromosomes,
alors que les Cercopithecidae ancestraux n'en
possédaient que 42. Ces évolutions en réseau s'opposent à
l'évolution dichotomique des lépilé-murs. L'existence d'un
nombre chromosomique élevé en cas d'évolution en réseau et
faible en cas d'évolution par dichotomie est caractéristique.
Le dernier groupe à considérer est celui des
Pongidae et des Hominidae . Un tronc commun les
individualise de l'ancêtre commun aux catarhiniens. Il est
probable que les Hylobatidae (gibbons) se soient
séparés assez tôt de ce tronc, mais les données cytogénétiques
sont peu informatives.
Revenons aux Pongidae pour constater que les
ancêtres de l'orang-outan se sont séparés d'abord. Le tronc
propre à l'orang-outan est court, et cette espèce possède deux
caryotypes distincts, qui correspondent aux formes de Suma-tra
et de Bornéo. Leur séparation géographique s'est donc
accompagnée d'une diversification chromosomique, ce qui
correspond vraisemblablement à un début de spéciation.
Malheureusement, l'extermination par l'homme de cette espèce
ne laisse aucune chance à cette évolution d'aboutir. Les
autres Pongidae et l'homme partagent ensuite un tronc
commun assez long, avant que leurs ancêtres ne se soient
séparés par une évolution en réseau. Du fait de la séparation
précoce de l'orang-outan et de la proximité du gorille et du
chimpanzé, il paraît clair que la branche des Hominidae
ne s'est séparée que très tardivement de la famille des
Pongidae . Nos ancêtres directs étaient donc bien des
Pongidae .
L'identification de notre plus proche parent a fait
couler beaucoup d'encre, tant de la part des scientifiques que
des philosophes, et des révélations sur le chaînon manquant
(lequel ?) sont régulièrement proposées. La seule originalité
de l'évolution chromosomique des Pongidae et des
Hominidae est de nous concerner de près.
D'une évolution en réseau sont issues les trois
branches menant aux espèces actuelles. Les derniers issus de
cette population commune sont vraisemblablement les ancêtres
des chimpanzés, puisqu'ils partagent les acquisitions du
gorille et de l'homme à la fois. Ils se trouvent donc en
position intermédiaire, et de ce fait plus près des ancêtres
des Hominidae . Ils sont aussi plus différenciés.
Les populations de gorilles, malheureusement bien
réduites aujourd'hui, ne semblent pas différer par leur
caryotype : celui du gorille des plaines est comme celui du
gorille des montagnes. La diversité est plus grande chez les
chimpanzés, où l'espèce commune ( Pan troglodytes ) et
l'espèce naine, aussi appelée bonobo ( Pan paniscus ),
diffèrent par deux modifications chromosomiques. Il est
possible qu'une autre modification différencie le chimpanzé «
blanc » du chimpanzé « noir*
», mais nous n'avons pu confirmer ce résultat, faute de
spécimens à étudier. En tout cas, les positions définies des
deux espèces de chimpanzés contre-disent formellement
certaines hypothèses récemment publiées selon lesquelles P.
paniscus serait un intermédiaire entre l'homme et le
chimpanzé commun.
Notre orgueil dusse-t-il en souffrir, il faut admettre
que la position de l'homme est bien banale dans cet arbre
évolutif. Nous sommes étonnamment proches des autres primates,
en particulier des Pongidae .
Nous souhaiterions terminer cette brève analyse qui
résume vingt-cinq ans de travaux sur une seule réflexion. Tout
biologiste rêve de connaître, autrement que par des fossiles,
les espèces qui ont jalonné l'histoire de la vie. Il est
difficile de pardonner aux hommes d'avoir fait disparaître
quelques-unes de ces espèces, par profit ou inconscience, et
surtout de continuer à le faire. L'homme est actuellement en
train d'éliminer de nombreuses espèces, et parmi les plus
menacées se trouvent l'orang-outan et le gorille, tellement
précieux pour la compréhension de notre propre histoire. C'est
une responsabilité insoutenable, qui sera jugée par nos
propres descendants comme l'un des pires crimes commis par
notre civilisation. A la différence des autres grands crimes
de ce siècle, il n'aura pas été commis par un dictateur fou,
mais par une humanité consciente. N'étant pas un enjeu
économique, il est malheureusement fort à craindre que cette
question ne soit pas traitée à temps au plus haut niveau
politique, à moins que même les mentalités puissent évoluer.
Bernard
Dutrillaux et Florence Richard 
Voir les références indiquées dans le numéro de mars
1997 de La Recherche . CYTOGÉNÉTIQUE
|
|
|