Analyse de la structure fige des chromosomes du Gorille (Gorilla gorilla)
Bernard Dutrillaux, Marie-Odile Réthoré, Marguerite Prieur et Jérôme Lejeune.
Humangenetik 20, 343-354 (1973).
Résumé :
Zusammenfassung. Prometaphase-Chromosomen von Gorilla, Mensch und schimpanse werden mit Hilfe der R-, Q-, T- und H-Bandentechnik verglichen; frühere Arbeiten (Lejeune et al., 1973) werden dadurch ergänzt. Verschiedene Mechanismen von Chromosomen-Rearrangements werden dargestellt; dabei finden die Heterochromatin-segmente besondere Beachtung. Einige phylogenetische Folgerungen werden gezogen.Sommaire
Dans un précédent travail (Lejeune et al.,1973), nous avons comparé la structure fine des chromatides de l'homme et du chimpanzé Pan troglodytes, à l'aide de quatre techniques de marquage.
Ces mêmes techniques sont utilisées pour l'étude du gorille (Gorilla gorilla) et les caryotypes des trois espèces sont ainsi comparés avec précision.
Un certain nombre de différences majeures avec les résultats de travaux antérieurs (Turleau et al.,1973 ; Grouchy et al., 1973 ; Pearson, 1973) sont mis en évidence.
Matériel et Méthodes
Les analyses chromosomiques ont été effectuées chez deux femelles, après culture de cellules sanguines, selon la méthode habituelle (Turpin et Lejeune, 1965 ; Dutrillaux et Couturier, 1972). Les quatre méthodes de marquage (bandes R, Q, T et H) précédemment utilisées pour la comparaison Pan-Homo (Lejeune et al., 1973) ont été employées.
Après l'établissement du caryotype de l'espèce, par l'analyse de métaphases en bandes R, nous avons cherché à reconnaître le maximum de structures par l'étude de prométaphases précoces, et comparé les résultats à ceux obtenus par les autres méthodes de marquage.
Résultats
Présentation des illustrations
Les fig. 1 à 5 montrent les chromosomes étudiés par les quatre techniques. A gauche est représenté un schéma de chromosome humain avec la nomenclature (Paris conférence, 1971). Ensuite, sur chaque ligne horizontale, on trouve deux images de dénaturation ménage (bandes R), puis deux images en bandes Q, puis deux images en bandes T et enfin deux images en bandes H. Dans chaque cas, on trouve à gauche un chromosome humain et à droite l'élément correspondant du gorille.
Bien que la comparaison ait porté sur les caryotypes de Homo, Gorilla et Pan, nous n'avons pas fait figurer les chromosomes de la dernière espèce, afin de ne pas alourdir les illustrations. On se référera donc au travail précédent (Lejeune et a1., 1973).
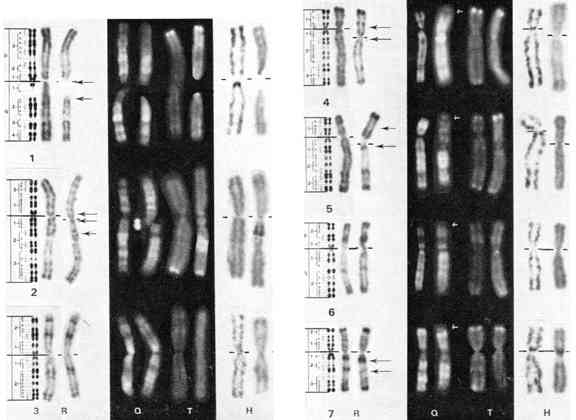
Fig. 1-5, Comparaison des chromosomes de l'homme et du
Gorille (Gorilla gorilla). De gauche à droite, analyse en bandes R, Q, T
et H. Dans chaque cas, le chromosome humain est à gauche et celui du
gorille à droite. Le schéma situe à l'extrême gauche représente la
nomenclature des chromosomes humains. Les flèches noires indiquent les
points de cassure des remaniements séparant les deux espèces ; les flèches
blanches, les bandes Q terminales surnuméraires chez le gorille.
Comparaison des chromosomes
La nomenclature des chromosomes humains est prise comme référence. Cependant, lorsque pour une paire donnée, les chromosomes du gorille sont identiques à ceux du chimpanzé, mais différents de ceux de l'homme, on indiquera simplement cette similitude sans donner le détail des remaniements chromosomiques qui a déjà été rapporté lors de la comparaison Pan-Homo (Lejeune et al., 1973).
Chromosomes 1
Bandes R, Q, T et H. Bras court et bras long identiques à ceux du chimpanzé.
Le gorille et le chimpanzé différent donc de l'homme par l'absence de la région q1 et des bandes q21 et q22.
Chromosomes 2
Comme chez le chimpanzé, deux éléments correspondent ici au chromosome 2 humain.
Chromosomes 2p
Plus strictement acrocentriques que chez le chimpanzé.
Bandes B et Q. Présence de deux bandes en plus sur le bras long et de deux en moins sur le bras court, ce qui peut indiquer qu'il s'est produit une inversion péricentrique entre Gorilla et Pan, avec des points de cassure situés en p11 et q12. Les satellites, probablement variables sont importants sur le bras court, alors qu'on ne les voit pas chez Pan.
Bandes T. Pas de marquage, comme chez Pan.
Bandes H. Marquage sur le bras court, qu'on observait mais moins important sur le bras long chez Pan, renforçant ainsi l'hypothèse d'une inversion péricentrique. Ce marquage a disparu chez l'homme.
Chromosomes 2q
Bandes R, Q, T et H. Identiques à celles du chimpanzé.
Chromosomes 3
Bandes R, Q, T et H. Identiques à celles de Pan et Homo.
Chromosomes 4
Ils ont un centromère moins distal que chez l'homme mais moins médian que chez le chimpanzé.
Bandes R. Par rapport à l'homme, bras court identique de p16 à p13 ; bras long identique de q35 à q21. I et inversion du segment p13 ? q13.
Il semble impossible de reproduire un chromosome 4 de chimpanzé à partir d'un 4 de gorille par un simple accident à deux cassures. II faudrait coït faire intervenir un accident à trois cassures (translation du segment q21.1 ? q21.2 dans la bande p13 du gorille, soit faire intervenir deux inversions péricentriques consécutives, le chromosome 4 humain représentant l'étape intermédiaire.
Bandes Q. Présence d'une bande Q terminale supplémentaire sur le bras court. Appellation proposée p17.
Présence d'une bande Q très fluorescente juxtacentromérique. Ce marquage, que l'on n'a jamais observé chez le chimpanzé, est beaucoup plus intense que celui qui existe, inconstamment, chez l'homme.
Bandes T. Marquage identique à Homo et Pan.
Bandes H. Absence de marquage, comme chez Pan.
Chromosomes 5
Ils ressemblent beaucoup à ceux du chimpanzé, mais en diffèrent par la partie terminale de leur bras court.
Bandes R. Par rapport à l'homme et au chimpanzé, bras long identique de q35 à q13.3. Le bras court rappelle celui du chimpanzé, mais son marquage diffère sensiblement, surtout à sa partie terminale.
Bandes Q. On n'observe pas la bande très fluorescente p14 existant chez Homo et Pan ; la fluorescence d'ensemble, du bras court, est relativement faible.
Présence d'une bande Q terminale supplémentaire chez Gorilla seulement.
Bandes T. Marquage terminal très important sur le bras court, sans correspondance chez Homo et Pan.
Bandes H. Absence de marquage.
Chromosomes 6
Bandes B, Q, T et H. Identiques à celles du chimpanzé, et différant seulement de l'homme par la présence d'une bande Q terminale sur le bras court.
Chromosomes 7
Bandes R. Bras court identique à celui de l'homme et du chimpanzé. Sur le bras long, la bande q11 est plus longue, et la bande q22 trop claire : possibilité d'une inversion paracentrique q11.3 ? q22.3
Bandes Q. Bras court : présence d'une bande Q terminale en plus de chez l'homme, comme chez le chimpanzé. Sur le bras long, rapprochement des deux bandes q21 et q31 du fait de la petite taille de la bande q22.
Bandes T. Identiques.
Bandes H. Identiques à celles du chimpanzé, c'est-à -dire, plus étendues que chez l'homme.
Chromosomes 8
Bandes R. Par rapport à l'homme et au chimpanzé, bras court identique de p23 à p21. Bras long identique de q24 à q22.1. Inversion péricentrique du segment p12 ? q21.3.
Bandes Q. Présence d'une bande Q terminale sur le bras court, comme chez le chimpanzé.
Bandes T. Identiques.
Bandes H. Pas de marquage, comme chez Homo et Pan.
Chromosomes 9
Bandes R. Bras long identique à celui de l'homme de q34 à q13. Inversion péricentrique du segment p24 ? q12. La bande q11, située sur le bras court chez Gorille, est plus marquée que chez Homo. Cette correspondance entre les bandes d'Homo et de Gorille ne se retrouve pas avec Pan où les premières bandes du bras long semblent d'une autre origine.
Bandes Q. Marquage juxtacentromérique présent chez Gorille seulement.
Bandes T. Identiques.
Bandes H. Le marquage observé sur le bras long du 9 humain peut fort bien correspondre à celui qui est observé ici sur le bras court.
On notera que la constriction secondaire du 9 humain équivaut ainsi au filament du satellite de son homologue chez le gorille.
Chromosomes 10
Bandes R. Par rapport à l'homme et au chimpanzé, bras long identique de q26 à q24. Inversion péricentrique du segment p15 ? q24 (fig. 3 et 6).
Bandes Q. Présence d'une bande Q terminale sur le bras "court", comme chez le chimpanzé.
Bandes T et H. Identiques.
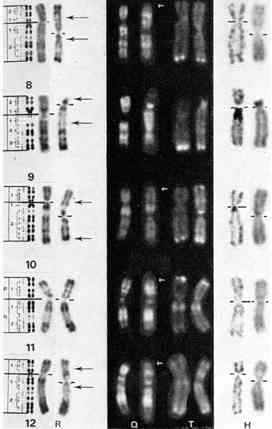
Fig. 6a-d. Chromosomes 10 métaphasiques. a Chromosomes 10
normaux du chimpanzé. b Chromosomes 10 humains normaux. c Chromosomes 10
d'une femme porteuse d'une inversion péricentrique : inv (11)) (p15; q24)
(élément de droite). d Chromosomes 10 normaux du gorille. Ces éléments
correspondent exactement au chromosome 10 remanié : inv (10) (p15; q24)
(Dutrillaux et al., 1973)
Chromosomes 11
Bandes. R, Q, T et H. Identiques à celles du chimpanzé, différant de celles de l'homme par la présence d'une bande Q terminale en plus, sur le bras court. Cette bande paraît moins intense chez Gorilla.
Chromosomes 12
Bandes R, Q, T et H. Identiques à celles du chimpanzé : inversion péricentrique et bande Q terminale en plus par rapport à l'homme.
Chromosomes 13
Bandes R, Q, T et H. Le bras long identique à celui de l'homme, diffère de celui du chimpanzé qui porte deux bandes intercalaires en plus. Bras court variable, mais généralement plus développé que chez l'homme, comme chez le chimpanzé. Seul le 13 humain porte un marquage Q juxtacentromérique.
Chromosomes 14
Bandes R et Q. Bras long identique à celui de l'homme et du chimpanzé, de q32 à q21. Inversion péricentrique du segment p11 ? q21. Les satellites du bras court, existant chez l'homme et le chimpanzé, sont ici généralement peu ou pas visibles.
Bandes T : Identiques.
Bandes H. Marquage de la partie juxtacentromérique du bras long, confirmant l'existence de l'inversion péricentrique.
Chromosomes 15
Bandes R, Q, T et H. Identiques à celles de l'homme : présence de deux bandes de plus que chez le chimpanzé.
Chromosomes 16
Bandes R. Par rapport à l'homme, bras court identique. Bras long identique de q24 à q13. La constriction secondaire, juxtacentromérique chez l'homme, est ici séparée du centromère par une bande sombre : inversion paracentrique possible du segment q11.1 ? q12. De plus, la constriction secondaire est en moyenne beaucoup plus développée chez le gorille.
Comme pour le chromosome 4, il ne paraît pas possible de reproduire le chromosome 16 de Pan à partir de celui de Gorilla, par un seul accident à deux cassures.
Il faudrait faire intervenir soit une translation de la constriction secondaire (trois cassures), soit deux inversions, péricentrique et paracentrique, et le 16 humain représenterait la forme intermédiaire.
Bandes Q. Bande Q terminale en plus, sur le bras court et le bras long, comme chez le chimpanzé.
Bandes T. Identiques.
Bandes H. Marquage de la constriction secondaire. Ce marquage existe aussi chez le chimpanzé, sur le bras court, mais fait défaut chez l'homme.
Chromosomes 17
Ils sont métacentriques, comme chez le chimpanzé, mais de marquage différent.
Bandes R. Le bras long peut correspondre à celui du chimpanzé, c'est-à -dire au segment q25 ? q21.3 de l'homme.
Le bras court, trop pâle, ne peut avoir la même origine que les 17 de l'homme et du chimpanzé.
Bandes Q. La bande très fluorescente, située au milieu du bras court, ressemble à celle que l'on observe sur le bras court du 5 de l'homme et du chimpanzé. L'hypothèse d'une translocation t(5;17) peut dans être envisagée. Présence d'une bande Q terminale sur le bras long.
Bandes T. Marquage limité à la bande terminale du bras long.
Bandes H. Pas de marquage, alors qu'on l'observe chez l'homme et le chimpanzé.
Chromosomes 18
Bandes R, Q, T et H. Identiques à celles du chimpanzé.
Par rapport à l'homme, inversion péricentrique et région hétérochromatique du bras court en plus.
Chromosomes 19
Bandes R et T. Identiques pour les trois espèces.
Bandes Q. Présence, comme chez le chimpanzé, d'une bande Q terminale en plus sur chaque bras.
Bandes H. Absence du marquage juxtacentromérique observé chez l'homme et le chimpanzé.
Chromosomes 20
Bandes R, T et H. Identiques pour les 3 espèces.
Bandes Q. Bande Q terminale sur chaque bras. Sur le bras long, cette bande est très importante, probablement double.
Chromosomes 21 et 22
Bandes R, T et H. Identiques pour les trois espèces.
Bandes Q. sur le bras long, présence d'une bande Q terminale en plus, comme chez le chimpanzé. Les bras courts, variables chez les trois espèces, sont généralement plus développés chez Gorilla et Pan.
Chromosomes X
Bandes R, Q, T et H. Identiques pour les trois espèces.
Chromosomes Y
Non étudiés ici.
Discussion
Seul le travail de Turleau et al. (1973) tentait de comparer les caryotypes de l'homme, du chimpanzé et du gorille, avec la méthode de bandes R.
L'analyse des stades prométaphasiques, et l'utilisation de plusieurs méthodes de marquage, apportent ici une précision sensiblement supérieure, et nos conclusions diffèrent sur la comparaison des chromosomes 2, 4, 5, 7, 9, 13, 14, 15, 16, 17 et 18.
a) Les chromosomes homologues
Comme nous l'avions signalé pour Pan et Homo (Lejeune et al., 1973), la méthode de bandes R s'avère la plus efficace pour la reconnaissance des similitudes chromosomiques.
Par cette méthode, on trouve huit homologies de marquage (chromosomes 3, 6, 11, 19, 20, 21, 22 et X) pour les trois espèces.
Si l'on tient compte des bandes Q, cette homologie ne persiste plus que pour les seuls chromosomes 3 et X, en raison des bandes Q terminales qui existent chez Pan et Gorilla, mais pas chez Homo.
Si l'on compare maintenant les espèces deux à deux, on reconnaît, en plus de ces homologies et en faisant abstraction de la variation des bandes Q terminales :
les 7, 8, 10 et 14 communs à Homo et Pan seulement,
les 13 et 15 communs à Homo et Gorilla seulement,
les 1, 12 et 18 communs à Pan et Gorilla seulement.
Au total, le caryotype humain se trouve plus proche de celui de Pan (12 homologies) que de celui de Gorilla (10 homologies), et le caryotype de Pan plus proche que celui d'Homo que de celui de Gorilla (11 homologies).
Les segments de chromatides homologues
Si l'on restreint l'analyse aux segments de chromatide, et non plus aux chromosomes entiers, les analogies recouvrent la presque totalité du caryotype.
Les seules structures dont il n'est pas possible de retrouver la correspondance avec certitude chez les trois espèces, sont les suivantes :
bandes 1q11 et 1q12, 1q21 et 1q22 présentes chez Homo seulement,
bandes 5p14 et 5p15, présentes chez Homo et Pan seulement,
bandes 9q12 et 9q13, reconnues chez Homo et Gorilla seulement,
deux bandes de 13q en plus, et deux bandes de 15q en moins, chez Pan seulement,
enfin, les trois bandes correspondant au 17p humain ne correspondent pas à celles du 17p de Gorilla.
L'absence d'homologie détectable ne porte donc que sur dix bandes environ entre Homo et Pan, Homo et Gorilla et Pan et Gorille. De plus, ce nombre relativement faible est peut être une surestimation de la réalité, puisque nous n'avons retenu que les homologies les plus évidentes.
Au total, si l'on considère que le caryotype de chaque espèce comprend prés de 500 structures, l'homologie est voisine de 98%.
D'autre part, il faut remarquer que les pertes ou gains de segments de chromatide ne sont pas plus importants entre Homo et Gorilla qu'entre Homo et Pan.
Les effets "quantitatifs" semblent donc avoir, dans l'évolution chromosomique d'espèces proches, un rôle mains important que les effets "cloisonnants" provenant de remaniements équilibrés.
c) Les remaniements de structure chromosomique
Les inversions péricentriques
Comme l'ont montré Turleau et al. (1973) et comme nous l'avons confirmé (Lejeune et al., 1973), les inversions péricentriques semblent avoir joué un rôle essentiel. On en dénombre :
huit, portant sur les chromosomes 4, 5, 8, 9, 10, 12, 14 et 18, et peut-être une neuvième portant sur le 17, qui séparent Homo de Gorilla,
six, portant sur les 2p, 4, 8, 10, 14 et 16, qui séparent Gorilla de Pan,
six, portant sur les 4, 5, 12, 16, 17 et 18, qui séparent Homo de Pan (Lejeune et al.,1973).
Il est intéressant de signaler que l'inversion péricentrique du 10, séparant Homo de Gorilla, se trouve observée dans un cas pathologique, chez l'homme (fig. 6), où elle est à l'origine d'un trouble méiotique par aneusomie de recombinaison.
Les inversions paracentriques
Ce type de réarrangement chromosomique n'avait pas été observé entre Pan et Homo. Bien qu'il soit difficile de l'affirmer, en raison de la petite taille des segments de chromatide remaniés, il semble qu'une inversion paracentrique se soit produite sur les chromosomes 7 et 16.
Pour le chromosome 7, une inversion du segment q11.3 ? q22.3 permet assez bien de reconstituer un chromosome de gorille à partir de celui de l'homme au du chimpanzé.
Pour le 16, une inversion du segment q11.1 ? q12 permet d'obtenir le chromosome du gorille à partir de celui de l'homme.
Ces inversions paracentriques pourraient avoir, à côté du rôle "cloisonnant", un rôle d'emblée "modifiant", du fait de la modification du sens de la lecture des gènes situés dans le segment inversé (Lejeune et al., 1973).
Les translocations
Lors de la comparaison Pan-Homo (Lejeune et ad., 1973), nous n'avions observé avec certitude qu'une seule translocation. Encore était-elle d'un type particulier puisque la fusion télomérique de deux éléments permettait d'obtenir le 2 humain. Aucune translocation de ce type ne sépare Gorilla de Pan et seule une éventuelle translocation entre un 5 et un 17 peut être discutée.
Si l'on compare maintenant les trois espèces, on constate que pour former le 2 humain, il faut d'abord "passer" de Gorilla à Pan. par une inversion péricentrique du 2p, la translocation s'effectuant nécessairement sur les mêmes chromosomes 2p et 2q que ceux du chimpanzé.
Les chromosomes 13 et 15 sont identiques chez le gorille et l'homme, alors que chez le chimpanzé, il manque deux bandes sur le 15 et qu'il y en a deux de plus sur le 13.
On peut donc supposer que les 13 et 15 observés chez l'homme et le gorille sont les plus proches de ceux de l'ancêtre commun. Il n'est pas exclu, d'autre part, qu'il se soit produit une insertion de deux bandes du 15 dans le 13 pour obtenir les chromosomes du chimpanzé.
Enfin, il semble qu'il existe une translocation t (5 ; 17).
Cependant, il est impossible de reproduire exactement les chromosomes 5 et 17 du gorille à partir de ceux de l'homme ou du chimpanzé, plusieurs remaniements seraient nécessaires.
d) Les bandes Q terminales
Nous avions vu, lors de la comparaison Pan-Homo, qu'il existait un grand nombre de bandes Q terminales, qui ne s'observaient que chez Pan.
D'une façon générale, ces mêmes bandes se retrouvent chez Gorilla. Seules différences, il existe une bande Q terminale sur le 5p, et pas sur le 17p chez Gorilla, ce qui pourrait confirmer l'hypothèse de la translocation t (5; 17) séparant les deux espèces. De plus, il existe une bande Q terminale sur le 4p de Gorilla, et pas chez Pan.
Au total, par rapport à Pan, il existe chez Gorilla une bande Q terminale en plus.
e) Les bandes Q juxtacentromériques
Il existe souvent, chez l'homme, un marquage Q intense, juxtacentromérique, des chromosomes 3, 4 et 13 (Paris conférence, 1971).
Lors de notre analyse de Pan, nous n'avions pas observé un tel marquage, mais devant le petit nombre d'animaux examinés, son absence systématique n'avait pu être affirmée.
Les mêmes remarques peuvent être faites pour les chromosomes 3 et 13 de Gorilla. Par contre, sur les chromosomes 4 de cette espèce, nous avons régulière ment observé un marquage très fluorescent. La forte intensité et la grande extension de ce marquage le distinguent nettement de celui du 4 humain.
Enfin, le chromosome 9, acrocentrique chez le gorille, porte un marquage qui n'a de correspondance ni chez Homo, ni chez Pan.
f) Les bandes T
Comme nous l'avions remarqué lors de la comparaison Pan-Homo, les bandes T sont très semblables d'une espèce à l'autre et semblent être des constituants chromosomiques très stables.
Seule différence avec Homo et Pan, la bande T du bras court du 5 est très forte, chez Gorilla, et son origine n'est actuellement pas connue.
g) Les bras courts des acrocentriques, les constrictions secondaires et les bandes H
1. Le chromosome 2p. Chez Gorilla, cet élément est acrocentrique et porte un marquage H sur son bras court. Chez Pan, il est submétacentrique et porte un marquage H sur son bras long.
2. Le, chromosome 9. Cet élément, chez Gorilla et chez Pongo (analyse en cours), est acrocentrique. Il porte, sur son bras court, du matériel hétérochromatique correspondant au satellite et à son filament. Cette région se colore fortement en bandes H.
Il paraît alors très probable que le chromosome de l'ancêtre commun ait été acrocentrique, et portait comme les autres éléments acracentriques, du matériel hétérochrornatique sur son bras court. Une inversion péricentrique permet d'obtenir le 9 humain, submétacentrique, dont la constriction secondaire et son marquage H particulier correspondrait alors Í l'hétérachromatine du bras court de l'acrocentrique d'origine.
3. Le chromosome 14. Chez Homo et Pan, cet élément est acrocentrique et porte un marquage H sur son bras court. Le même chromosome 14 est retrouvé chez Pongo (analyse en cours). Par contre, chez Gorilla, le 14 est submétacentrique et porte un marquage H sur son bras long.
Bien qu'il soit encore trop tôt pour en tirer une conclusion générale, il apparaît donc, comme le montre la formation du chromosome 9 humain, du 2p du chimpanzé et du 14 du gorille, que lorsqu'un élément non acrocentrique possède un marquage H, ce marquage provient de l'hétérochromatine du bras court d'un acrocentrique ancestral. Il sera donc très intéressant de connaître l'origine des autres chromosomes non acrocentriques possédant, eux aussi, un marquage H.
Un certain nombre d'autres remarques méritent d'are faites à propos des bandes H :
toutes sont situées à proximité d'un centromère,
tous les acrocentriques portent ces bandes dont les dimensions sont assez variables,
chez une même espèce, elles semblent fréquemment subir de petits remaniements, comme en témoigne l'inversion péricentrique du 9 souvent observée chez l'homme,
les chromosomes portant ces structures semblent subir de fréquents remaniements, au cours de l'évolution, et ces faits sont à rapprocher de la pathologie humaine où, par exemple, les translocations atteignent si souvent les acrocentriques,
lorsqu'elles sont éloignées du centromère, à la suite d'un remaniement, elles semblent perdre leurs propriétés tinctoriales particulières,
enfin, ce marquage qui semble relativement indépendant des autres, peut "apparaître" ou "disparaître", sur un chromosome donné, sans modifier les marquages R et Q de même localisation.
Ainsi, une simple analyse morphologique de ces structures laisse supposer qu'elles possédent une constitution chimique particulière, prédisposant peut-être aux remaniements chromosomiques.
Conclusions
L'analyse du caryotype du gorille confirme et précise les conclusions que nous avions portées, lors de la comparaison Pan-Homo, sur les mécanismes de l'évolution chromosomique chez les primates hominiens.
1. Les remaniements de type "cloisonnant" par inversion péricentrique en particulier, restent, par leur fréquence, le mécanisme le plus apparent.
2. Les changements "quantitatifs" tels que les additions et les délétions de segments de chromatides ont peut être joué un rôle plus limité, n'étant pas plus importants entre Homo et Gorilla qu'entre Homo et Pan.
3. Les accidents chromosomiques observés en pathologie humaine et les changements chromosomiques séparant les trois espèces ne semblent pas fortuits et peuvent relever des mêmes facteurs favorisants. La constitution de l'hétérochromatine marquée en bandes H est peut être l'un de ces facteurs.
4. Entre Gorilla et Pan, on n'observe pas de translocation par fusion télomérique dont le rôle, à la fois "cloisonnant" et "modifiant" avait été discuté lors de la comparaison Pan-Homo (Lejeune et al., 1973).
Par contre, l'inversion paracentrique du 7, et peut être du 16, séparant Gorilla d'Homo et Pan, pourrait jouer le même rôle.
5. Bien que l'étude phylogénique complète ne sera entreprise qu'ultérieurement, après l'analyse du caryotype de Pongo, un certain nombre de drainons peuvent déjà être mis en évidence.
Par exemple, pour reproduire le chromosome 2 humain à partir des 2p et 2q de Gorilla, il est indispensable de passer par l'étape intermédiaire représentée par les 2p et 2q de Pan.
Inversement, le chromosome 4 humain semble représenter l'étape intermédiaire entre les 4 de Pan et de Gorilla. En effet, alors qu'on passe simplement d'Homo à Gorilla, ou d'Homo à Pan par une simple inversion péricentrique, il n'est pas possible de passer du 4 de Pan à celui de Gorilla à moins d'un remaniement exigeant un minimum de trois cassures. Il en est peut être de même pour le chromosome 16.
Bibliographie
Dutrillaux, B., Couturier, J.: Techniques d'analyse chromosomique. In. Biologie génétique. Paris: l'Expansion 1972
Dutrillaux, B., Laurent, C., Robert, J. M., Lejeune, J.: Inversion péricentrique, inv (10), chez la mère et aneusomie de recombinaison, inv. (19), rec (10), chez son fils. Cytogenet. 12, 245-253 (1973)
Grouchy, J. de, Turleau, C., Roubin, u., Chavin Colin, F.: Chromosomal evolution of man and the primate (Pan troglodytes, Gorilla gorilla, Pongo pygmaeus). In : Proc. Nobel Symp.1972. Chromosome Identification, Technique and Applications in biology and medicine. Nobel Foundation Stockholm. New York-London: Academic Press 1973
Lejeune, J., Dutrillaux, B., Réthoré, M. O., Prieur, M.: Comparaison de la structure fine des chromatides d'Homo sapiens et de Pan troglodytes. Chromosoma (Berl.) 43, 423-444 (1973)
Paris Conference: 1971 Standardization in human cytogenetics. The National Foundation, March of Dimes, Vol. VIII Nb 7 (1972)
Pearson, P.: The uniqueness of the human karyotype. In: Proc. Nobel Symp. 1972. Chromosome Identification, Technique and Applications in biology and medicine. Nobel Foundation Stockholm. New York-London: Academic Press 1973
Turleau, C., Grouchy, J. de, Klein, M.: Phylogénie chromosomique de l'homme et des primates hominiens (Pan troglodytes, Gorilla et Pongo pygmaeus). Essai de reconstitution du caryotype de l'ancêtre commun. Ann. Génét. 15, 225-240 (1972)
Turpin, R., Lejeune, J.: Les chromosomes humains. Paris: Gauthier Villars 1965