 |
|
01/03/1997 La
Recherche
BERNARD
DUTRILLAUX Institut Curie et département de Radiobiologie
et Radiopathologie DSV/CEA.
 Figure
1. Figure
1.
 Figure
2. Figure
2.
 Figure
3. Figure
3.
 Figure
4. Figure
4.
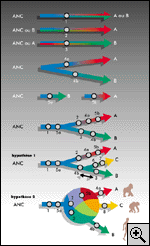 Figure 5. Figure 5.
*HÉTÉROZYGOTE De hétéro (différent) et
zygote (oeuf fécondé). Employé ici au sens d'un
individu dont les deux allèles (copies provenant de l'un et
l'autre parent) d'un même gène sont différents.
*BANDES Depuis 1970, diverses techniques de
coloration des chromosomes font apparaître des bandes
caractéristiques, qui permettent d'en préciser la morphologie
et de procéder à des comparaisons systématiques.
*PHÉNOTYPE Désigne ce qui résulte de
l'expression du génotype (ensemble de gènes). L'apparence d'un
individu constitue donc un phénotype.
*SÉGRÉGATION CHROMOSOMIQUE Séparation et
répartition des chromosomes pendant la méiose.
*PROPHASE Premier stade de la division
cellulaire. Dans le cas de la méiose, les chromosomes
s'apparient puis recombinent.
*THEODOSIUS DOBZHANSKY Généticien russe
installé aux Etats-Unis après 1940, auteur d'une série
d'ouvrages fondamentaux sur l'évolution.

(1) M.J.D. White, Modes
of speciation , Editions Freeman and Co., San Francisco,
1978.
(2) S. Jay Gould et N. Eldredge, Nature, 366
, 223, 1993.
(3) D. Petit, B. Dutrillaux, Ann. Génét.,
28 , 13, 1985.
(4) Y. Rumpler et B. Dutrillaux, «
Chromosomal evolution and speciation in primates », Cell
Biology Reviews , juin 1990, Springer.
(5) B.
Dutrillaux, Ann. génét., 29 , 69, 1986.
(6) Th.
Dobzhansky, Genetics of the evolutionary process ,
Columbia University Press, 1970.
(7) S. Ohno, Evolution
by gene duplication , Allen and Unwin, Londres, 1970.
La Recherche a publié
:
B. Dutrillaux, « Les chromosomes des primates »,
novembre 1981.
| |
|
|
|
> DOSSIER
>
CYTOGÉNÉTIQUE
Les remaniements chromosomiques
jouent un rôle majeur dans la spéciation
Comment évoluent les
chromosomes des mammifères
L'analyse des caryotypes de quelque
deux cents espèces et sous-espèces de mammifères, primates
compris, révèle des surprises. Les remaniements chromosomiques
sont fréquents, mais ne s'accumulent pas au même rythme selon
les groupes. Chez les espèces qui possèdent le plus grand
nombre de chromosomes, le modèle classique de l'évolution par
bipartition, fondé sur l'étude de la drosophile, n'est pas
applicable. Il faut lui substituer un modèle en réseau.
L'apparition d'une espèce
nouvelle est un événement fondamental et encore mal appréhendé
par les théoriciens de l'évolution. L'approche traditionnelle
consiste à faire intervenir la conjonction de deux facteurs :
une série de mutations, plus ou moins aléatoires au niveau
moléculaire, et la pression de la sélection naturelle,
favorisée par l'isolement géographique ou écologique. Je pense
pour ma part que les remaniements chromosomiques jouent un
rôle majeur, souvent mésestimé, dans la mécanique évolutive,
et en particulier dans la spéciation.
Lorsque deux animaux ou groupes d'animaux diffèrent par
certains remaniements chromosomiques, ils ne peuvent pas se
reproduire durant plusieurs générations, car la fabrication
des gamètes ne peut se faire convenablement chez les
hétérozygotes*. Ces remaniements créent donc une barrière, la
barrière gamétique, qui peut devenir une barrière d'espèce.
Les remaniements chromosomiques sont des phénomènes
fréquents (fig.
2). Chez les mammifères, leur analyse
a beaucoup progressé ces dernières années. Dans notre
laboratoire, nous avons comparé les chromosomes de plus de
deux cents espèces et sous-espèces de mammifères : primates,
carnivores et rongeurs.
On a cru longtemps que chaque espèce avait son propre
caryotype. Ce n'est pas totalement exact. La comparaison de
nombreuses espèces montre que, dans certains groupes, des
espèces pourtant proches morphologiquement ont des caryotypes
très différents, alors que dans d'autres groupes, des espèces
bien distinctes morphologiquement possèdent des caryotypes
proches ou identiques. J'entends par même caryotype un même
nombre de chromosomes, avec les mê-mes bandes*
et, aussi loin qu'on aille dans l'analyse de localisation des
gènes, les mêmes gènes situés au même endroit. La carte
génétique ne montre donc pas de différence. Toutefois, on
trouverait des différences au niveau des gènes, à la suite des
mutations qui ont pu survenir.
L'existence de ces situations surprenantes montre que
dans certains grou- pes les modifications des chromosomes
s'accumulent vite par rapport au rythme d'accumulation des
mutations ponctuelles à l'intérieur des gènes. De ce fait, le
phénotype*, en particulier la morphologie, évolue peu et le
caryotype beaucoup, tandis que dans d'autres groupes c'est
l'inverse, le caryotype reste relativement stable, mais la
morphologie varie beaucoup, en raison d'un taux de mutations
géniques élevé. Cela relativise la signification de l'horloge
biologique, dont on parle beaucoup : elle marche à des
vitesses différentes selon les groupes et les paramètres
choisis. Une horloge qui marche à des vitesses différentes
n'est donc pas une bonne horloge !
Concernant le rôle des modifications chromosomiques,
les scientifiques ne se sont guère intéressés, jusqu'à
présent, qu'à l'effet immédiat des remaniements chromosomiques
: le rôle de barrière gamétique(1). Pourquoi ? Parce qu'on
sait, notamment à partir de la pathologie humaine, qu'un
hétérozygote porteur d'un tel remaniement a une descendance
qui tend à être plus anormale que celle de la moyenne de la
population. La reproduction de l'hétérozygote étant
défavorisée, on en conclut que l'évolution devait fonctionner
de façon très particulière pour expliquer l'acquisition d'un
remaniement : le passage par une toute petite population,
voire par un couple unique, comme le conçoit le mythe d'Adam
et Eve. On n'imagine pas d'autre solution qu'un croisement
entre hétérozygotes, même si cela entraîne des difficultés de
procréation, pour qu'au hasard des ségrégations*
chromosomiques se forme une population homozygote pour ce
remaniement.
L'idée qu'une nouvelle espèce puisse émerger par un tel
processus est paradoxale puisqu'il a toujours été admis que
l'évolution privilégie des situations favorables, dans le
cadre de la sélection naturelle. Comment alors expliquer la
survenue fréquente des remaniements chromosomiques, au cours
de l'évolution, alors qu'être porteur hétérozygote d'un tel
remaniement défavorise la procréation ?
En réalité, on a entretenu une confusion entre l'effet
d'un remaniement du caryotype sur le phénotype et l'effet de
ce même remaniement sur la reproduction. Un individu peut être
porteur d'une particularité génétique ou chromosomique et, en
raison d'une dérive dans la fabrication des gamètes, fabriquer
plus de gamètes d'un type que d'un autre. Qu'une telle dérive
existe et il est alors possible d'avoir plus de descendants
porteurs de la particularité génétique, bien que celle-ci
entraîne un certain nombre de difficultés de procréation.
A l'échelle de l'évolution, les individus ne sont que
les vecteurs d'une information génétique, et il ne faut pas
confondre les difficultés qu'ils ont éventuellement à vivre
avec leur information génétique, et la difficulté, qui peut
être bien moindre, de transmettre cette information. Une
translocation susceptible de créer une descendance à
problèmes, si elle est associée à un gène favorable à la
reproduction, peut finalement se transmettre au point de créer
une population nouvelle. Un désavantage génétique immédiat
peut donc être largement compensé, sur le long terme, par un
avantage associé.
On a aussi beaucoup exagéré le rôle défavorable des
remaniements chromosomiques pour la descendance, en raison
d'un double préjugé. D'abord, nous avons transféré sur les
autres espèces ce que nous savions de la nôtre. Il est vrai
qu'il n'existe pas de population humaine, plus ou moins
isolée, porteuse d'un remaniement chromosomique particulier.
Notre espèce est essentiellement monomorphe : les
translocations ou inversions récurrentes ont des fréquences
inférieures à 1 ä. De ce fait, chaque variant, chaque porteur
d'un remaniement, est considéré comme potentiellement
pathologique. Cette situation ne s'applique manifestement pas
à beaucoup d'autres espèces, où l'on observe un polymorphisme
chromosomique. Dans ces espèces, le fait d'être porteur d'un
remaniement chromosomique ne peut pas être considéré comme
pathologique.
Le second préjugé résulte d'un biais d'analyse. On ne
fait pas l'examen chromosomique de tout le monde. Il faut un
motif, et ce motif est le plus souvent l'existence d'une
anomalie visible. C'est le biais de recrutement des individus,
par la médecine, qui a amené à exagérer le rôle défavorable
des remaniements. Or, il existe dans la population des
individus porteurs de translocations ou d'inversions
chromosomiques, qui pourtant se reproduisent parfaitement
bien, sans difficulté apparente.
On sait même que dans certains cas des inversions
portées par des hommes normaux reconstituent des chromo-somes
de chimpanzé ou de gorille. Elles n'ont donc pas d'effet sur
le phénotype. Cela montre aussi que ces inversions ne se
produisent pas tout à fait au hasard. Elles se font en des
points privilégiés, probablement en raison d'homologies de
séquences favorisant la recombinaison de l'ADN.
Il ne faut pas oublier que l'immense majorité de notre
génome est constitué de séquences dont on ne connaît pas la
fonction, qui ne sont pas des gènes, en ce sens qu'elles ne
codent pas pour une protéine donnée. Il existe donc une
infinité de ruptures possibles de chromosomes sans rupture de
l'information génétique. Il y a d'ailleurs des raisons de
penser qu'il existe beaucoup plus de remaniements entre
séquences non codantes qu'entre séquences codantes.
N'oublions pas non plus que ce que nous observons dans
la population est le résultat d'une forte sélection in
utero . Beaucoup d'embryons sont éliminés très tôt en
raison de déséquilibres chromo-somiques, qui peuvent être
induits par des remaniements.
A partir du moment où l'on admet qu'un remaniement
chromosomique peut être à l'origine d'une spéciation, il est
tentant, on l'a vu, d'admettre l'idée d'un goulot
d'étranglement originel, couple unique ou micropopulation.
Stephen Jay Gould étaye en partie sa théorie des équilibres
ponctués sur ce type d'hypothèse : une espèce se formerait
rapidement après la survenue de mutations « spécifiques(2) ».
Je n'y crois pas beaucoup.
L'étude des mammifères, en particulier des primates et
surtout des rongeurs, montre qu'il existe des populations
polymorphes. Ce polymorphisme semble très bien supporté, avec
des hétérozygotes et des homozygotes pour diverses formules
chromosomiques.
On ne peut donc ni affirmer que le caryotype de la
plupart des espèces soit homogène, ni supposer que ce
polymorphisme pénalise fortement la reproduction des
hétérozygotes. En fait, il ne faut pas considérer l'effet d'un
seul remaniement chromosomique, mais de plusieurs. Le
polymorphisme chromosomique permet une accumulation
progressive de remaniements dans une population. Par le biais
des croisements entre individus, il est envisageable de voir
apparaître, au bout d'un temps plus ou moins long, des
individus porteurs homozygotes de plusieurs remaniements
chromosomi-ques susceptibles de générer une barrière
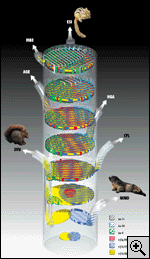
gamétique(3) (fig.
1).
Ce processus peut être comparé à la survenue des
maladies récessives : une mutation génétique se transmet de
manière neutre dans la population, jusqu'au jour où deux
personnes porteuses de la même mutation se rencontrent et
procréent. Dès lors, des homozygotes peuvent naître porteurs
du caractère anormal ou nouveau.
L'évolution chromosomique procède à mes yeux de cette
façon, créant une sorte de réseau. Des remaniements
chromosomiques s'installent dans la population, s'il n'y a pas
de forte contre-sélection, permettant l'instauration d'un
polymorphisme chromosomique. Cela ne suffit pas, bien sûr, à
créer une espèce nouvelle. Mais l'effet des remaniements peut
être comparé à celui d'un isolement géographique. Ils
individualisent une micro-population qui a de plus en plus de
mal à se reproduire facilement avec le reste de la population
d'origine.
Autre fait important : deux espèces proches possèdent
souvent un même chromosome impliqué dans deux remaniements
différents chez l'une et l'autre(4). Ce point nous a longtemps
échappé, mais c'est probablement un élément important du
cloisonnement qui se crée entre les descendants. En effet, le
polymorphisme qu'on observe dans les populations montre que
des individus peuvent être hétérozygotes pour plus d'un
remaniement et se reproduire quand même. En revanche aucun
exemple n'est connu de reproduction d'individu porteur de deux
remaniements affectant deux chromosomes homologues. Cela
semble entraîner des troubles considérables de la méiose.
Quittons maintenant l'effet immédiat pour aborder un
point qui a été jusqu'ici complètement négligé : les
remaniements des chromosomes peuvent avoir un effet à long
terme, sur la spéciation, sur des durées significatives à
l'échelle de l'évo-lution.
Si l'on considère l'arbre évolutif des primates, tel
qu'il a pu être reconstitué, on s'aperçoit que chez les
prosimiens (les lémurs) la grande majorité des remaniements,
environ neuf sur dix, sont des translocations par fusion de
deux chromosomes (fig. 2). Au cours de l'évolution des
pongidés et de l'homme, neuf fois sur dix ce sont des
inversions qui surviennent, alors que chez les cercopithèques
ce sont le plus souvent des fissions, entraînant une
augmentation du nom-bre des chromosomes. Cette accumulation de
modifications chromosomiques n'est donc pas aléatoire.
Pourquoi ? Il n'existe encore aucune explication
satisfaisante, mais une interprétation semble s'imposer dans
certains cas au moins. Je pense que l'évolution, d'une façon
générale, procède par étapes et que chaque étape prédétermine
les suivantes. Cela signifie simplement que les mutations
orientent la réponse à la sélection, et que par la suite la
sélection orientera le succès des autres mutations.
Un exemple nous est fourni par les translocations entre
le chromosome X et un chromosome non sexuel (autosome). Chez
l'homme, de telles translocations conduisent toujours à une
réduction de fertilité ou à des malformations, tandis que chez
certains rongeurs, par exemple dans la famille des gerbilles,
les remaniements entre chromosomes sexuels (gonosomes) et
autosomes sont très fréquents.
Comment expliquer cette différence ? Le chromosome X
est très particulier. Il y en a un seul chez les mâles et deux
chez les femelles, dont un est inactivé. Les gènes réglant
cette inactivation sont maintenant connus. Cette inactivation
s'applique à la quasi-totalité des gènes portés par le
chromosome. Les études de pathologie humaine nous disent que
chez la femme, lorsqu'un autosome est attaché à un X inactivé,
l'inactivation peut se propager sur une partie au moins de
l'autosome. Ce chromosome perd sa fonction, et l'individu ne
survit pas, ou est malformé.
Chez l'homme, une translocation de l'X, ou de l'Y, ne
retentit pas sur le phénotype. Par contre, dans les cellules
germinales, pendant la prophase de la méiose, quand l'X et l'Y
sont inactivés tandis que les autosomes sont transcrits, une
translocation autosome-gonosome se traduira par une
inactivation de l'autosome qui subit la translocation, un
arrêt de la gamétogenèse, et finalement une stérilité. Les
seules exceptions concernent des remaniements où des séquences
d'ADN hautement répété se retrouvent intercalées entre le
gonosome et l'autosome. Dans ce cas, l'inactivation ne se
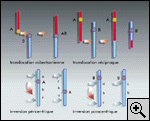
propage pas du gonosome à l'autosome (fig.
3). Or, dans toutes les espèces où surviennent des
translocations X-autosome, le segment de l'autosome est séparé
du segment du chromosome X par une zone d'ADN très particulier
(hétérochromatine constitutive) qui porte justement des
séquences d'ADN très répété. On sait que l'X est généralement
métacentrique chez les mammifères : c'est-à-dire que le
centromère, constriction du bâtonnet chromosomique, est situé
au milieu de celui-ci. Mais chez les gerbilles ancestrales,
l'X a subi une inversion péricentrique (fig. 2) qui l'a rendu
acrocentrique : le centromère, associé à de l'ADN très répété,
s'est retrouvé à une extrémité du chromosome, qui ensuite, a
pu fusionner avec tout autosome, le segment autosomique se
trouvant séparé du segment gonosomique.
Un processus à deux temps est donc bien survenu : le
remaniement qui amène de l'hétérochromatine à l'extrémité d'un
chromosome sexuel rend possible sa translocation ultérieure
avec un autosome. Bien que cet exemple soit unique, je présume
que ce type d'événement survient souvent. La difficulté est
qu'on ne sait encore presque rien des structures
chromosomiques qui ne sont pas les gènes, et qui jouent
vraisemblablement un rôle dans la dynamique chromosomique.
Je crois à une évolution à tiroirs, où une première
mutation rend le génome, et donc l'individu, ouvert à de
nouvelles modifications, qui auparavant étaient délétères.
Force est en tout cas de constater que, du fait de
cette accumulation non aléatoire des remaniements, les
formu-les chromosomiques changent consi-dérablement. Chez les
lémurs, l'accumulation de translocations aboutit à une
réduction progressive du nombre de chromosomes, jusqu'à
seulement 20 chromosomes chez certains Indridae . Les
fissions qui surviennent chez les cercopithèques aboutissent
au résultat inverse : les ancêtres des cercopithèques avaient
autour de 44 chromosomes, tandis que certaines espèces en ont
aujourd'hui jusqu'à 72.
Nous avons reconstitué le caryotype ancestral des
primates. Il comportait une soixantaine de chromosomes, tous
composés d'un bras et d'un centromère à une extrémité
(acrocentrique), à l'exception de l'X. L'accumulation
d'inversions permet de déplacer le centromère vers le centre,
de sorte que chaque chromosome porte deux bras séparés par le
centromère. Ainsi, le nombre de bras peut doubler sans
augmentation du nombre de chromosomes.
Pour résumer, le caractère non aléatoire des
remaniements chromosomi-ques fait que certains types
s'accumulent. De ce fait, le nombre de chromosomes et le
nombre de bras chromosomiques d'un groupe donné devient très
différent de ceux du groupe voisin.
Sauf exception, une espèce ne se forme pas à partir
d'une seule modification génétique. En général, la comparaison
de deux espèces voisines montre qu'elles sont séparées par
quelques pour- cent de différences géniques. Evidemment, on ne
sait pas exactement ce qui s'est passé au départ, mais l'on
suppose qu'il faut plus d'une mutation pour instaurer un
phénotype correspondant à une nouvelle espèce. Supposons, pour
simplifier, qu'il faille trois mutations pour que des
individus porteurs se reconnaissent à la fois semblables entre
eux et différents des autres. Si ces trois mutations sont
proches les unes des autres sur un même chromosome, elles vont
se transmettre souvent comme un seul gène, tout en codant
chacune pour un caractère précis. A l'inverse, si elles sont
portées par trois chromosomes différents, du fait de la
ségrégation indépendante des chromosomes à la méiose, elles
vont être redistribuées à chaque génération avec les divers
chromosomes. Le groupe de caractères ne pourra donc se
maintenir. Dans ce cas, la spéciation ne pourra se faire
d'emblée. Un polymorphisme se développera dans la population,
et c'est seulement au hasard de croisements plus ou moins
éloignés dans le temps que les trois mutations pourront
devenir plus fréquentes dans une sous-population, et
finalement se retrouver ensemble dans un groupe d'individus.
Le nombre de chromosomes constituant le caryotype de
l'espèce va donc jouer un rôle considérable. Une espèce avec
un nombre très faible de chromosomes aura une forte
probabilité de transmettre ensemble les trois mutations. Très
vite, à l'échelle de l'évolution, se détachera un rameau
constituant une spéciation par bipartition, c'est-à-dire par
dichotomie.
En revanche, une telle évolution est impossible pour
une espèce avec un grand nombre de chromosomes puisque les
mutations sont continuellement redistribuées au hasard des
ségrégations chromosomiques. Celles-ci peuvent être simplement
évaluées par la formule S = 2N où N est le nombre haploïde (en
une seule copie) de chromosomes.
Ainsi, dans la famille des Cercopithecidae , le
nombre de combinaisons possibles est infiniment plus grand
chez certains mones (cercopithèques d'Afrique-Equatoriale), où
N = 36, que chez les babouins et macaques où N = 21(5).
Jusqu'ici, nous avons fait abstraction des
recombinaisons entre chromosomes homologues, qui surviennent
au stade prophase*
de la méiose. Ces recombinaisons font que chaque chromosome
transmis dans les gamètes est une mosaïque de la paire de
chromosomes homologues d'origine : c'est le brassage génique
par recombinaison ou crossing-over. Ces crossing-over laissent
une trace : les chiasmas, dénombrables au microscope au stade
métaphase de la première division méiotique. Il est donc
possible de comparer leur nombre chez des espèces à caryotypes
différents. Ce décompte nous a permis de montrer qu'il existe
au moins un chiasma par bras chromosomique, donc au moins un
par chromosome acrocentrique (le centromère étant à une
extrémité, il n'y a qu'un seul bras) et deux par chromosome
métacentrique, quelle que soit leur taille. En conséquence, il
existe une forte relation entre le nombre de chromosomes, et,
mieux encore, de bras chromosomiques, et le taux de
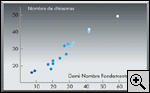
recombinaison méiotique (fig.
4). En d'autres termes, deux gènes situés à même
distance physique sur un même segment chromosomique auront
d'autant moins de chances d'être recombinés que ce segment est
situé sur un grand chromosome.
L'évolution chromosomique non aléatoire, qui a
progressivement entraîné des fragmentations dans certains
taxons, ou inversement des fusions de chromosomes dans
d'autres, a donc considérablement modifié les probabilités de
transmission de groupes de mutations. Il en résulte que les
taxons à nombre réduit de chromosomes pourront évoluer sur un
mode dichotomique, alors que ceux possédant un grand nombre de
chromosomes ne pourront évoluer qu'en réseau. Les dynamiques
d'apparition de nouvelles espèces s'en trouveront changées.
Dans l'évolution dichotomique, le phénomène de
spéciation pourra être relativement régulier. A l'opposé, dans
l'évolution en réseau, ou populationnelle, elle se fera par
à-coups. Il faudra attendre que les mutations se disséminent,
entraînant un polymorphisme intraspécifique, qu'elles se
répartissent diversement dans la population, et qu'enfin
apparaissent des homozygotes pour des mutations différentes en
des points séparés de ladite population. A une période de
latence succédera donc une période d'explosion où plusieurs
espèces s'individualiseront à partir d'une même population
d'origine (voir fig. 1 et 5).
Un exemple typique est fourni par les cercopithèques
arboricoles. Issus d'une même population ancestrale, des
groupes plurispécifiques se forment encore aujourd'hui et des
éthologues ont même remarqué l'existence d'un mâle dominant à
phénotype hybride, ce qui peut contribuer fortement à
redistribuer les mutations, donc à une dynamique en réseau.
Ce schéma évolutif n'est pas admis classiquement.
N'oublions pas que les théories modernes de l'évolution et de
la génétique des populations sont en partie basées sur les
travaux de l'école de Dobzhansky*,
qui décrivent de belles évolutions dichotomiques chez les
drosophiles(6). Or les drosophiles sont des diptères possédant
8 chromosomes (4 pai-res) seulement. De plus, il n'y a pas de
recombinaison méiotique chez le mâle. Toutes les conditions
sont donc réunies pour faciliter la transmission en bloc de
groupes de mutation, et donc une évolution dichotomique. Ce
modèle ne peut donc être généralisé, et s'applique mal aux
mammifères, dont la très grande majorité des espèces possède
entre 40 et 60 chromosomes.
Le dernier point que je souhaite souligner est la
nature non aléatoire des mutations géniques. Là encore, une
école de pensée dogmatique a introduit la notion que les
mutations étaient aléatoires, et conféré du coup un rôle quasi
exclusif à la sélection qui ne retient que les bonnes
mutations.
Avec l'accumulation des données sur la nature des
mutations, cette vision paraît simpliste. Les notions de
dominance (la mutation d'un allèle sur deux s'exprime) et de
récessivité (les deux allèles doivent être mutés pour que le
caractère mutant s'exprime) gardent une forte signification.
Il apparaît que la grande majorité des mutations induites par
les mutagènes, environnementaux ou autres, sont récessives, et
qu'elles entraînent une perte de fonction de l'allèle touché,
l'autre allèle s'exprimant seul. De telles mutations peuvent
s'accumuler, mais ne sont peut-être pas très intéressantes
pour la dynamique de l'évolution, puisqu'elles orientent vers
des pertes de fonction. Elles entraîneraient plutôt le
vieillissement d'une population en alourdissant son fardeau
génétique. Ce processus est peut-être l'explication du grand
mystère qu'est la disparition quasi-systématique des espèces
ancestrales.
Les mutations dominantes semblent plus intéressantes,
puisqu'elles créent une nouvelle fonction. Elles semblent
survenir rarement et sont peu induites par les mutagènes. Deux
cas de figure existent.
Dans le premier cas, une fonction se substitue à une
autre, en passant par la phase hétérozygote. Le plus souvent,
un phénotype remplacera un autre, mais il n'y aura pas de
grand changement, en particulier de gain de fonction. Ces
mutations jouent vraisemblablement un rôle dans le
polymorphisme du phénotype, comme la variation de la
pigmentation et de caractères morphologiques ou immunologiques
mineurs.
Le second cas de figure aboutirait au gain de fonction.
Dans la mesure où tout laisse supposer que les gènes en place
ont leur utilité, il paraît intéressant de créer de nouveaux
gènes vecteurs de nouvelles fonctions. L'existence de la
duplication de certains gènes a été suspectée depuis
longtemps(7), elle est maintenant démontrée. Les remaniements
chromosomiques semblent bien jouer, là aussi, un rôle en
permettant la transposition d'un gène d'un chromosome à
l'autre, ou d'une position à l'autre d'un même chromosome.
Ainsi apparaît une structure nouvelle, directement dérivée
d'un gène existant qui reste en place. Il peut s'agir d'une
simple duplication, qui aura l'intérêt de permettre par une
mutation ultérieure l'acquisition d'une nouvelle fonction,
sans perte de la fonction originale. Il peut s'agir encore de
la formation d'un gène tronqué, qui formera un pseudogène non
transcrit, mais qui constituera un réservoir de matériel
génétique, pour d'éventuelles mutations futures.
Pour résumer, les réarrangements chromosomiques
semblent jouer plusieurs rôles essentiels dans la dynamique de
l'évolution :
barrière gamétique, à la manière de l'isolement
géographique ;
orientation vers une évolution dichotomique ou en
réseau (populationnelle) ;
enfin moteur dans l'acquisition de nouvelles
fonctions potentielles. Il est donc regrettable que leur rôle
et leur importance soient trop souvent négligés.
Bernard
Dutrillaux 
« Evolution chromosomique chez les prima-tes, les
carnivores et les rongeurs », Mammalia 50 ,
numéro spécial, 1986. SPECIAL : L'HISTOIRE DE LA VIE
CYTOGÉNÉTIQUE
|
|
|




